 Back to Resources
Back to Resources Introduction
Since the mid-1980s fertility of UK dairy herds has declined year-on-year To put this into perspective, in 2014 the University of Nottingham reported that poor fertility was estimated to cost the UK dairy industry £350 million annually. This included lost milk output, excessive culling and increased veterinary costs. It is argued that genetic selection for high milk production has compromised fertility. Although this might be true to a limited extent, poor fertility can be attributed to many factors including genetic, nutritional, and environmental and management.
Within the dairy industry, statistics show a number of dairy farms achieve excellent milk yield and fertility records. A host of factors revolving around nutrition and management are largely responsible for the variations in performance between dairy enterprises.
In nutrition, inadequate supply of the right balance of essential fatty acids could be a causative factor to poor fertility but this has received little recognition at farm level. This is because traditionally fat has been considered singularly as an energy source with disregard to its other vital role in cow health and fertility. It is no wonder when ration formulation programmes allocate no values to constituent fatty acids as opposed to protein where the contribution to milk yield of individual amino acids are considered. It is intriguing that the decline in cow fertility accelerated to some degree with the withdrawal of fishmeal from dairy rations. Fish oils are rich in omega-3 fatty acids which have been implicated in improving reproductive performance in animals (Silvestre et. al, 2011a).
Role of fats in dairy cow nutrition
Fat has traditionally been fed to dairy cows because its calorific content is higher than that of the other feed ingredients and it therefore increases the energy density of the total diet. However, beyond the supply of energy for production and growth, constituent fatty acids also impact on health and reproduction of the dairy cow. Furthermore, because of human health concerns, the fatty acid profile of dairy products is becoming increasingly influential on consumer choice. Therefore, consideration of fat composition is important not only in formulating dairy rations but also with regard to human health.
Particular fatty acids are considered essential either because the animal cannot synthesise them or they only produces them with limited efficiency. They include the polyunsaturated fatty acids (PUFA), linoleic acid (LA), and alpha linolenic acid (ALA). Eicosapentaenoic acid (EPA) and Docosahexaenoic acid (DHA) which are found mainly in fish oils and marine algae are also considered essential. This is because they can be produced from ALA but the efficiency of conversion is variable at best and very low at worst.
PUFA are the building blocks of phospholipids in cell membranes and influence structural integrity and function of body cells. They are precursors of lipid mediator molecules and as such are involved in neurological development, health and reproduction processes in the animal. For example, it has long been known that EPA and DHA can suppress production of prostaglandins which may influence early embryonic mortality in dairy cows. At the same time they increase levels of progesterone, the pregnancy maintenance hormone (Thatcher et al., 1994 and Mattos et. al., 2004). ALA is only 30% as effective (Mattos et al., 2003). Both Ω3 fatty acids also exhibit anti-inflammatory properties and play a crucial role in the post-calving recovery process.
Most recent work by researchers at University of Florida (Greco, 2015) demonstrated that feeding a diet with a ratio of Ω6 to Ω3 of 4:1 increased dry matter intake with concurrent improvement in milk yield, butterfat, protein and lactose and resulted in lower somatic cell count. There also seems to be a synergistic effect of that combination of fatty acids on energy yield.
In the dairy cow, rumen biohydrogenation is a major impediment to the delivery and absorption of the essential fatty acids. In their unprotected form, more than 80% of dietary PUFA are modified by rumen fermentation and therefore not available to the animal in their original form. The challenge is how to ensure adequate supplies of these essential nutrients in order to cater for the dairy cow’s own requirement for optimal performance and provide for the consumer health needs via the dairy products.
Fat supplementation and limitations in dairy rations
Presently, the two common commercial methods of rumen protection are either the formation of calcium salts of palm fatty acids or the hydrogenation of fats to render them insoluble in rumen. The resistance to ruminal and intestinal lipolysis is a major factor contributing to poor digestibility of hydrogenated fats which reduces their potential value in dairy cow rations. According to Sukhija and Palmquist (1990), the most abundant calcium salt of palm fatty acid distillate requires a minimum rumen pH value of 5.5 to remain protected from dissociation. However, such a rumen environment is difficult to maintain at all times with high concentrate diets designed for high yielding herds.
Due to the high melting point, hydrogenated fats may not be readily digested and absorbed in the lower gut. With regard to calcium salt formation, the technique works well with saturated and medium chain fatty acids but it has not been successful with PUFA due to the high pH threshold value of 6.3 necessary to avoid dissociation in the rumen (Van Nevel and Demeyer, 1996). The calcium salts of long chain fatty acids are therefore not stable at normal rumen pH. Moreover, such products have fixed formulations focused on supplying energy and increasing butterfat levels rather than meeting the cow’s other needs for essential fatty acids for health and reproduction. They also have no influence on milk fat composition.
Fat supplements high in C16 can drive up milk fat and loss in body condition, both of which impact negatively on cow health and fertility (Leroy et. al., 2013). It can therefore be argued that the leading brands of supplemental fats may not be the most appropriate vehicle for delivering the essential fatty acids to the dairy cow and producing healthier milk.
New approach to fat supplementation
The contest is to make fat supplements available with the capability of delivering the right balance of the essential fatty acids for intestinal absorption and tissue uptake without ruminal moderation. Feeding vegetable oils either in a straight form of as oil seeds has been tried but they suffer the drawback of milk fat depression and reducing dry matter intake (Givens et. al., 2009). Fortunately, fat supplements are now available that deliver essential fatty acids to the dairy cow without being altered in the rumen or affecting butter fat levels. These blends of vegetable and marine oils are encapsulated within the fibre matrix of the carrier base.
A trial carried out at CEDAR, Reading University on behalf of UFAC-UK showed that matrix protection of vegetable and marine oils using cellulolytic fibre as a carrier was effective within three hours of incubation in the rumen. We believe this is achievable with our method of rumen protection based on small particles with specific gravity equal to that of the rumen fluid. The carrier particle size is ≤2mm which is below the threshold of 2 – 4 mm for rumen passage in cattle. On absorption of water, the products are able to attain the specific gravity identical to that of the rumen fluid necessary for rumen escape. These two factors, together with high feed intake typical of high yielding cows, would facilitate rapid escape of the products from the rumen before any significant degree of biohydrogenation takes place. It has also been reported (Wasowska, et. al., 2009) that fish oils inhibit ruminal biohydrogenation partly by a mechanism that affects activity of B. fibrisolvens which is the principal biohydrogenating bacterium.
Recently, UFAC-UK conducted an observational trial to verify the findings at Reading University and validate the effectiveness of fibre matrix protection using the products dynalac and Promega. dynalac is a 50% dry fat product consisting of vegetable and marine oils while Promega is a fishmeal replacer based on vegetable proteins and fish oil. 400g and 300g of each respectively were fed per cow per day to a well-managed UK milking herd averaging 32 litres in early lactation. Both products have higher levels of unsaturated than saturated fatty acids (combined fatty acid composition is shown in table 1).
We have compared our milk fat profile with the average milk fat composition from cows fed 0-800g/cow/day protected fat (Nottingham University, 2002-2005) and another milk fatty acid profile outcome from a study carried out on the fatty acid make-up of winter retail milk by the University of Newcastle (see table 2 and fig 1 ).
Table 1. Fatty acid profile of dynalac and Promega.
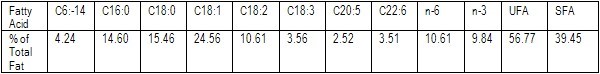
Table 2. Comparison of milk fat profiles.

The University of Nottingham study carried out between 2002 and 2005 was funded by DEFRA and involved varying levels of protected fat (0 – 800g/cow/day) and starch in dairy cow diet and determining their influence on milk fat composition.
The winter trial was conducted in North East England by the University of Newcastle (Butler, et. al., 2011) over a two year period (2006-2008) and involved sampling of 12 different brands of milk produced under conventional winter feeding systems. The objective of the study was to identify any variations in fatty acid composition of UK retail milk.
Although our data has not been generated from a scientifically designed experiment some notable differences emerge when we compare the composition of milk fat from the different sources which provides food for thought.
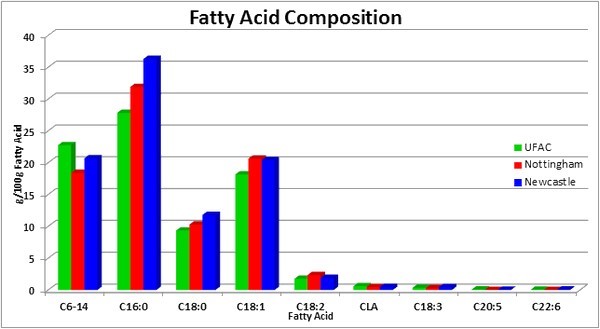
Overall the observation confirms our conviction that the carrier matrix within which the oils are physically entrapped in our products provides an effective means of protection from rumen biohydrogenation. This is shown by the significantly lower level (10.2%) of saturated fat in milk produced from our fat supplement-based rations compared with that from the conventional winter diet and 5.75% less than the industry average of 69% (table 2).
On the other hand, the content in milk fat of the short chain fatty acids is higher (23.6%) in our diet compared with those of the protected fat, which is an indication of better rumen function and healthier cows. It is also important to note that under conditions of negative energy balance, concentrations of short chain fatty acids decrease (Palmquist, et. al. 1993).
The level of C16:0 is 12.8% lower in milk from our herd when rated against the milk fat from the cows fed protected fat. About 50% of the C16:0 in milk originates from blood having come directly from the diet or from the adipose tissue following body fat mobilisation. As such, there is a positive correlation between the increases in the level of C16:0 in milk fat and negative energy balance.
The level of C18:1 in our diet is 11.6% lower compared with the other observations. High content of this fatty acid in milk would signify body fat mobilisation. There is also emerging evidence that lameness is associated with breakdown of adipose tissue which depletes the digital fat pad that cushions the feet from the pressure from the cow’s body weight. The fatty tissue in the digital pad consists mainly of monosaturated fatty acids of which C18:1 is a major component. The prevalence of lameness in dairy herds in England and Wales stands at around 36.8% and should be a warning to the dairy industry (Barker, 2010).
There does not seem to be any difference among the treatments on the level of stearic acid in the milk fat. If anything, the fatty acid profile from our diets shows less C18:0. Strearic is the major end product of complete rumen biohydrogenation.
The relatively higher level of conjugated linoleic acid (CLA) in milk from our herd is significant because biomedical studies have shown that this fatty acid has important health benefits including anti-carcinogenic and anti-atherogenic activities. CLA is also known to alter nutrient partitioning away from body fat and to boost immune function. While some CLA may be produced as an intermediate in the rumen bioyhdrogenation of linoleic acid, this is only a transient as 80% of CLA originates from endogenous synthesis in the mammary gland.
The ratio of omega 6 to omega 3 fatty acids in the milk fat from our diet is similar to that of conventional winter feeding but greater than that of protected fat and indeed closer to the ideal ratio of 4:1. In the recent past researchers (Greco et. al., 2014) in United States have reported improved milk yield and fertility by manipulating the dietary fatty acids to achieve this ratio. Specifically pregnancy rate increased from 30% to 37% and pregnancy loss declined to 6% from 12%. There was also an increase in feed efficiency, measured as kgFCM/kgDM fed of 5-6%, superior anti-inflammatory response and a reduction in somatic cell count. Body condition loss was also minimised.
The Omega-3 fatty acids (EPA and DHA) content in our milk is 0.12% compared with none in the protected fat ration. Omega-3 fatty acids are of great interest because of the potential benefits to human health. The effects of these fatty acids in cardiovascular disease, type II diabetes, hypertension, cancer plus certain neurological functions have been well documented by Calder (2013). Within the cow the Omega-3 fatty acids have positive influences on health and fertility.
EPA and DHA are generally absent or present at minimal levels (<0.1% of total fat), in common dairy diets and as a consequence are lacking in milk and dairy products. Even with the use of omega-3 rich feedingstuffs in dairy cow rations only modest increases in the levels in milk have been achieved. This is because efficiency of transfer of these particular dietary fatty acids into milk fat is low, averaging 3.0% and 2.0% for EPA and DHA respectively (Chilliard, et. al. 2001). Moreover, the Omega-3 fatty acids are also required by other target tissues in the adipose as well as reproductive, immune and neurological systems. However, when fish oils are administered post-ruminally or fed in rumen-protected-form transfer efficiencies have been increased substantially with 32% for EPA and 18% for DHA being recorded for formaldehyde-protected tuna oil.
In our case the cows produced 32 litres of milk at 3.79% fat received a daily ration supplying 12.5g of EPA and DHA from marine oils which were rumen-protected within the matrix of vegetable fibre carrier. Based on the average milk fat, and the 0.12% content of total fat for both fatty acids, the combined yield was 1.45g. This represents almost 12% transfer efficiency which is more than a threefold increase over what has been observed with unprotected forms of fish oils, bearing in mind that these fatty acids are also deposited elsewhere in other tissues within the body. The omega-3 content is similar to that observed by US researchers when calcium salts of sunflower and fish oils were fed to achieve the omega 6 to omega-3 ratio of 4:1, which resulted in improvement in milk yield, immune response and reproductive performance.
Sustainable cow productivity
Although for many farmers the immediate economic agenda is seeking to maximise revenue through higher levels of bufferfat, the health and fertility of the cow need to be taken into consideration for long term benefits. This objective can significantly be achieved through feeding fat supplements carefully designed to optimise cow’s productivity, health and fertility.
Stakeholders in the dairy industry including nutritionists, producers, processors and consumers need to work together in providing for the economic interest of the farmer, improved cow welfare and consumer health. Key to this will be reversing the decline in cow fertility and producing healthy milk through rationalised nutritional and management interventions.
Conclusion
- The fibre matrix method of protecting oils from rumen biohydrogenation provides a means of delivering the essential fatty acids to the target tissues of the animal for better health and fertility. This allows formulation of dairy cow diets for improved reproduction, rather just for increased milk production.
- The implications of being able to manipulate dietary fats and influence milk fat composition for the dairy industry are significant, with the prospect of introducing feeding and breeding programmes based on products that will improve cow health and also increase the nutritional value of milk within the human diet.
- There is a benefit in using milk fatty acid profiling as a tool to assess the nutritional, health and fertility status of dairy herds.
- The short-term gains from producing milk with higher levels of fat should not undermine the long term beneficial effects of proper dietary rationing on cow health and fertility.
- Feeding fat supplements high in essential fatty acids including omega-3s offers economic benefit to the farmer in terms of sustainable herd productivity, and heathier dairy products to the consumer.
- The dairy industry needs to take advantage of the ability to influence milk fatty profile through dietary manipulation to add nutritional value to dairy products and counter pressure for their reduction in intake.
References
Barker Z. E., Leach K. A., Whay H. R., Bell N. J., Main D. C. J.( 2010). Assessment of lameness prevalence and associated risk factors in dairy herds in England and Wales. J. Dairy Sci. 93, 932-941.
Butler G., Stergiadis S., Seal C., Eyre, M. and Leifert, C. (2011). Fat composition of organic and conventional retail milk in north east England. Dairy Sci. 2011 Jan; 94 (1):24-36. doi: 10.3168/jds.2010-3331
Calder, P. C. (2013). Omega-3 fatty acids in disease prevention: a general overview. http://www.nutri-facts.org/eng/expert-opinion/detail/backPid/93/article/omega-3-fatty-acids-in-disease-prevention-a-general-overview/
Chilliard, Y., Ferlay, A., and Doreau, M. (2001) Effect of Different Types of Forages, Animal Fat or Marine Oils in Cow’s Diet on Milk fat Secretion and Composition, Especially Conjugated Linoleic Acid (CLA) and Polyunsaturated Fatty Acids, Livest. Prod. Sci. 70, 31-48.
Givens, D. I., Kliem, K. E., Humphries, D. J., Shingfield, K. J. and Morgan, R. (2009) Effect of replacing calcium salts of palm oil distillate with rapeseed oil, milled or whole rapeseeds on milk fatty-acid composition in cows fed maize silage-based diets. Animal, 3:7, pp 1067–1074.
Greco, L. F., Neves Neto, J.T., Pedrico, A., Ferrazza, R. A., Lima, F.S., Bisinotto, R. S., Martinez, N., Garcia, M., Ribeiro, E. S., Gomes, G.C., Shin, J. H., Ballou, M. A., Thatcher, W.W., Staples, C.R. and Santos, J. E. (2015) Effects of altering the ratio of dietary n-6 to n-3 fatty acids on performance and inflammatory responses to a lipopolysaccharide challenge in lactating Holstein cows. J Dairy Sci. Jan;98(1):602-17. doi: 10.3168/jds.2014-8805. Epub 2014 Nov 14
Leroy, J.L.M.R., Sturmey, R.G., Hoeck, V. Van., De Bie, J., McKeegan, P.J. and Bols, P.E.J. Dietary lipid upplementation on cow reproductive performance and oocyte and embryo viability: a real benefit? (2013). Anim. Reprod., v.10, n.3, p.258-267, Jul. /Sept. 2013
Mattos, R., Guzeloglu, A., Badinga, L., Staples, C. R., Thatcher, W. W. (2003). Fatty Acid Regulation of Endometrial Prostaglandin Secretion. (http://www.biolreprod.org/content/early/2003/04/30/biolreprod.102.015057.full.pdf)
Mattos, R., Staples, C. R., Arteche, A. C. M., Wiltbank, M. C., Diaz, F. J., Jenkins, T. and Thatcher, W. W. (2004). The effects of feeding fish oil on uterine secretion of PGF2α, milk composition, and metabolic status of periparturient Holstein cows. J. Dairy Sci. 87:921-932
Palmquist, D.L., Baulieu, A.D. and Barbano, D.M. 1993. Feed and animal factors influencing milk fat composition. J. Dairy Sci. 76:1753-1771.
Silvestre, F.T., Carvalho, T.S., Francisco, N., Santos, J.E.P., Staples, C.R., Jenkins, T.C. and Thatcher, W.W. (2011a). Effects of differential supplementation of fatty acids during the peripartum and breeding periods of Holstein cows: I. Uterine and metabolic responses, reproduction, and lactation. J. Dairy Sci. 94:189-204.
Sukhija, P. S. and Palmquist, D. L. (1990).Dissociation of calcium soaps of long-chain fatty acids in rumen fluid. J Dairy Sci. Jul;73 (7):1784-7.
Thatcher, W. W., Staples, C. R., Danet-Desnoyers, G., Oldick, B. and Schmitt, E. P. (1994). Embryo health and mortality in sheep and cattle. J. Anim. Sci. 72 Suppl.3:16 -30.
University of Nottingham, School of Biosciences (2002-2005). Nutritional, Hormonal and Genetic Influences on Milk Fat Composition in Dairy Cows.DEFRA SID 5 Research Project Final Report.
Wasowska, I., Maia, M. R., Neidzweidzka, K. M., Czauderna, M., Ribeiro, J. M., Devillard, E., Shingfield, K. J. and Wallace, R. J. (2006). Influence of fish oil on ruminal biohydrogenation of C18 unsaturated fatty acids. Br J Nutr. Jun;95(6): 1199-211.
Van Nevel, C. J. and Demeryer D.I. (1996). (Effect of pH on biohydrogenation of polyunsaturated fatty acids and their Ca-salts by rumen microorganisms in vitro. Arch Tierernahr. 1996;49(2):151-7
For further information please contact our technical department on 01780460327 or technical@ufacuk.com